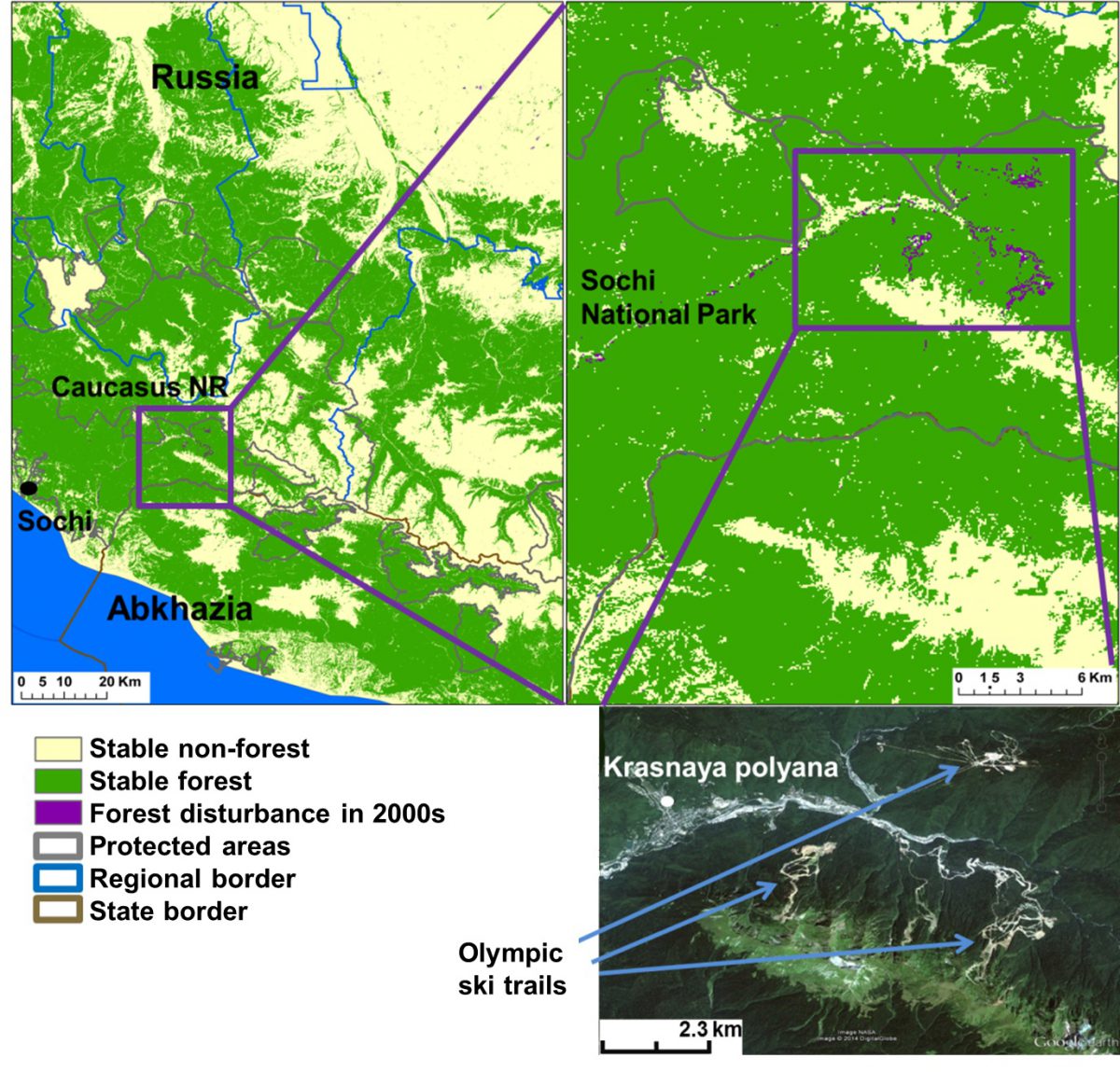
Spatial Analysis For Conservation and Sustainability
Biodiversity
Species richness varies by orders of magnitude across the globe and so does the richness of endangered species. We are interested in both the macroecological processes causing richness patterns, and the socioeconomic processes determining where biodiversity is threatened.
The lack of management experience at the landscape scale and the limited feasibility of experiment at this scale have increased the use of scenario modeling to analyze the effects of different management actions on focal species. However, current modeling approaches are poorly suited for the analysis of viability in dynamic landscapes. Demographic (e.g., metapopulation) models of species living in these landscapes do not incorporate the variability in spatial patterns of early successional habitats, and landscape models have not been linked to population viability models. We link a landscape model to a metapopulation model and demonstrate the use of this model by analyzing the effect of forest management options on the viability of the Sharp-tailed Grouse ( Tympanuchus phasianellus) in the Pine Barrens region of northwestern Wisconsin (U.S.A.). This approach allows viability analysis based on landscape dynamics brought about by processes such as succession, disturbances, and silviculture. The landscape component of the model (LANDIS) predicts forest landscape dynamics in the form of a time series of raster maps. We combined these maps into a time series of patch structures, which formed the dynamic spatial structure of the metapopulation component (RAMAS). Our results showed that the viability of Sharp-tailed Grouse was sensitive to landscape dynamics and demographic variables such as fecundity and mortality. Ignoring the landscape dynamics gave overly optimistic results, and results based only on landscape dynamics (ignoring demography) lead to a different ranking of the management options than the ranking based on the more realistic model incorporating both landscape and demographic dynamics. Thus, models of species in dynamic landscapes must consider habitat and population dynamics simultaneously.
Management of German roe deer (Capreolus capreolus) populations is a challenge for wildlife managers and foresters because population densities are difficult to estimate in forests and forest regeneration can be negatively affected when roe deer density is high. We describe a model to determine deer population densities compatible with forest management goals, and to assess harvest rates necessary to maintain desired deer densities. A geographic information system (GIS) was used to model wildlife habitat and population dynamics over time. Our model interactively incorporates knowledge of ?eld biologists and foresters via a graphical user interface (GUI). Calibration of the model with deer damage maps allowed us to evaluate density dependence of a roe deer population. Incorporation of local knowledge into temporally dynamic and spatial models increases understanding of population dynamics and improves wildlife management.
Habitat quality is an important consideration when identifying source and sink habitat and setting priority areas for avian conservation. The problem is that different measures may lead to different conclusions about habitat quality, and may also vary in the resources required to estimate them. Individual level measures, such as nest success, and fecundity, will often identify different high quality habitats than population level measures, such as abundance or the number of fledglings produced per unit area. We tested measures of fitness in the Black-throated Sparrow both at the individual and at the population level for six habitats in the northern Chihuahuan Desert, to explore their value as indicators of habitat quality. We compared clutch size, number of nestlings per nest, number of fledglings per successful nest, nest density, nest success, daily nest survival rate, season-long fecundity, number of fledglings produced per 100 ha, and adult abundance, in each habitat type. We also modeled source-sink dynamics to estimate the scale at which they operate, to infer survival rates, and to ascertain the relative source potential of each habitat. We found that fecundity is the best indicator of individual level habitat quality but a poor indicator of population level habitat quality. Nest success (or fecundity, if resources are available to adequately estimate it) plus nest density provide the most robust indicator of population level habitat quality, which is the level at which priority habitats for conservation should be identified. Mesa grassland and black grama grassland functioned as source habitats most consistently, and mesquite was consistently a sink but also probably a reservoir of individuals available to occupy other habitats.
1. Where populations are con?ned to fragmented, human-dominated landscapes, preventing declines and extirpations will often rely on metapopulation management. Spatially-explicit population viability analyses provide tools to evaluate how well the local management e?orts can be combined to conserve metapopulations across large areas. Yet, metapopulation models have rarely been combined with tools to assess the cost-e?ectiveness of di?erent conservation strategies. 2. European bison Bison bonasus only occur in small, fragmented populations, making their long-term survival dependent on establishing a metapopulation across eastern Europe. We parameterized a European bison metapopulation model based on time-series of bison demography and a habitat suitability map to assess the viability of bison populations in the Carpathians and the relative cost-e?ectiveness of (i) reintroductions, (ii) wildlife overpasses and (iii) anti-poaching measures in establishing a viable bison metapopulation. 3. Our results suggest that the Carpathians could support a viable metapopulation of European bison provided that active e?orts are taken to safeguard bison and connect isolated herds. With such steps, our model forecasts that bison numbers could increase substantially over the next 100 years as local populations increase and bison recolonize parts of the Carpathians. 4. Reintroductions appear to be the most cost-e?ective approach for establishing a viable bison metapopulation among our scenarios, especially when coupled with wildlife overpasses to improve connectivity among herds. The most promising region for a bison metapopulation in the Carpathians was south-eastern Poland, Ukraine and northern Romania. We identi?ed several candidate regions for reintroductions and wildlife overpasses, especially in the border region of Romania and Ukraine. Site-speci?c assessments of both habitat suitability, and the costs and bene?ts of a large bison population, should target those regions. 5. Synthesis and applications. Our results highlight how careful conservation planning can identify solutions to preserve large mammals in human-dominated landscapes. Choosing the most e?ective option from a range of management strategies is a central challenge for wildlife managers. We have shown that incorporating cost-e?ectiveness analyses into metapopulation models can elucidate the relative value (gain per unit cost) of di?erent conservation management options, allowing decision makers to choose cost-e?ective options to preserve large mammals. Our model projections also provide hope for establishing a viable free-ranging European bison population in the Carpathians, one of the last relatively wild areas in Europe.
Habitat loss threatens large mammals worldwide and their survival will depend on habitat in human-dominated landscapes. Conservation planners thus face the challenge to identify areas of least conflict with land use, yet broad-scale species distribution models rarely incorporate real landscape patters nor do they identify potential conservation conflicts. An excellent example of such conservation challenges are European bison. Almost extinct by the early 20th century, bison can only survive in the wild if large metapopulations are established, but it is unclear where new herds can be reintroduced. Using European bison as an example we conducted a continental-scale habitat assessment based on real landscape patterns. Our specific aims here were to a) map European bison habitat throughout the species' former range, b) examine whether broad-scale habitat suitability factors differ from previously reported fine-scale factors, and c) assess where suitable habitat occurs in areas with low potential for conflict with land use. We assessed habitat suitability using herd range maps for all 36 free-ranging European bison herds as habitat use data. Habitat suitability maps were compared with maps of land cover, livestock density, agricultural constraints, and protected areas to assess potential conservation conflicts. Our models had high goodness-of-fit (AUC = 0.941) and we found abundant potential bison habitat. European bison prefer mosaic-type landscapes, with a preference for broadleaved and mixed forests. European bison metapopulations appear not limited by habitat availability. However, most potential habitat occurred outside protected areas and has substantial potential for conservation conflicts. The most promising areas to establish large bison metapopulations all occur in Eastern Europe (i.e., the Carpathians, the Belarus-Ukraine borderlands, and several regions in European Russia). The future of European bison and that of other large mammals in the wild thus clearly lies in Eastern Europe, because habitat there is most abundant and least fragmented, and because the potential for conflict with land use is lower. More generally we suggest that broad-scale habitat assessments that incorporate land use can be powerful tools for conservation planning and will be key if large herbivore and carnivore conservation is to succeed in a human-dominated world.
Understanding past and current patterns of species richness is essential for predicting how these patterns may be affected by future global change. The species energy hypothesis predicts that higher abundance and richness of animal species occur where available energy is higher and more consistently available. There is a wide range of remote sensing proxies for available energy, such as vegetation productivity, but it is not clear which best predict species richness. Our goal here was to evaluate different proxies for annual plant productivity from Terra and Aqua Moderate Resolution Imaging Spectroradiometer (MODIS) as input for the Dynamic Habitat Indices (DHIs), and to determine how well they predict the richness of breeding bird species in six functional guilds across the conterminous United States. The DHIs are measures of vegetation productivity over the course of a year and consist of three components: (1) cumulative productivity (DHI Cum), (2) minimum productivity (DHI Min), and (3) intra-annual variation of productivity (DHI Var). We hypothesized that increases in cumulative and minimum productivity and reductions in intra-annual variation will be associated with higher species richness. We calculated the DHIs from a range of MODIS 1000-m vegetation productivity data sets for 2003– 2014, i.e., the Normalized Difference Vegetation Index (NDVI), Enhanced Vegetation Index (EVI), Fraction of absorbed Photosynthetically Active Radiation (FPAR), Leaf Area Index (LAI), and Gross Primary Productivity (GPP). We summarized bird species richness of different guilds within ecoregions (n = 85) based on abundance maps derived from the N3000 routes of the North American Breeding Bird Survey for 2006 to 2012. Generally, we found all the DHIs had high explanatory power for predicting breeding bird species richness. However, the strength of the associations between the DHIs and bird species richness depended on habitat, nest placement, and migratory behavior. We found highest correlations for habitat-based guilds, such as grassland breeding species (R2 adj 0.66–0.73 for the multiple DHI regression model; R2 adj 0.41–0.61 for minimum DHI) and woodland breeding species (R2 adj 0.34–0.60 for the multiple DHI regression model; R2 adj 0.26–0.51 for cumulative DHI). The strong relationship between the DHIs and bird species richness reinforces the importance of vegetation productivity as a determinant of species diversity patterns, and the usefulness of satellite data for applying the species energy hypothesis to predictions in service to conservation.
Many animal and plant species found in areas where they did not occur in the past. These newcomers are known as invasive species. The consequences of having a new species could benefit the system by bringing new ecosystem services to the areas such as new pollinators for crops. Unfortunately in many cases the new species can have negative impacts at the community level by competing with local species and even displacing them. Diana is particularly interested in a species that has had mainly negative consequences for other species in places wherever it was introduced, the mongoose. Mongooses ( Herpestes auropunctatus) are native from India, and were introduced to Caribbean and Pacific islands at the end of the 20th century. These predators were brought into the area to control rats (another invasive) that were affecting sugarcane plantations. However, mongooses didn’t only eat rats it also local island species such as birds, amphibians and reptiles. Therefore this new species caused the decline and extinction of local species without eradicating the rat problem.
Mongoose in the live trap will be checked for disease
Local and federal agencies have tried to remove or reduce populations of mongoose from the islands but have failed because mongoose populations grew rapidly. Although they are omnivorous, mongooses keep predate especially local populations of ground nesting birds and marine tortoises among other species. However, there is little knowledge of the extent of the problem: how mongooses behave, its biology in the island, which species is eating the most or how the populations are related within and between islands.
Diana is studying mongooses in Puerto Rico from multiple angles, to have a bigger picture of the problem. She wants to know where the mongooses populations are and how well connected the populations are to each other. She considers that populations that are connected are more difficult to manage, because removed mongooses will be easily replaced by individuals of adjacent connected populations. Diana will use molecular techniques to identify these connection patterns and how are natural or artificial barriers and other habitat features in the landscape limiting mongoose dispersal. She also wants to know what are preferred prey species of different populations of mongoose. By using stable isotopes, she will track the source of food of mongooses.
Probability of mongoose occupying habitats in eastern Puerto Rico. Connectivity prediction was done using 34 mongooses trapped on the eastern part of the island via CIRCUITSCAPE using structural resistance. (Data on Cabo Rojo NWR was excluded)
Ultimately, Diana will have a sense of how the overall island community of wildlife species is assembled, having this predator already stablished from over a century. This information will be useful for wildlife managers and biologist who will know how the mongoose is distributed and prioritize areas to control the populations. With information about the trophic position and links of this predator, she will provide the base for research questions related to novel animal community assemblies and resilience after species introductions.
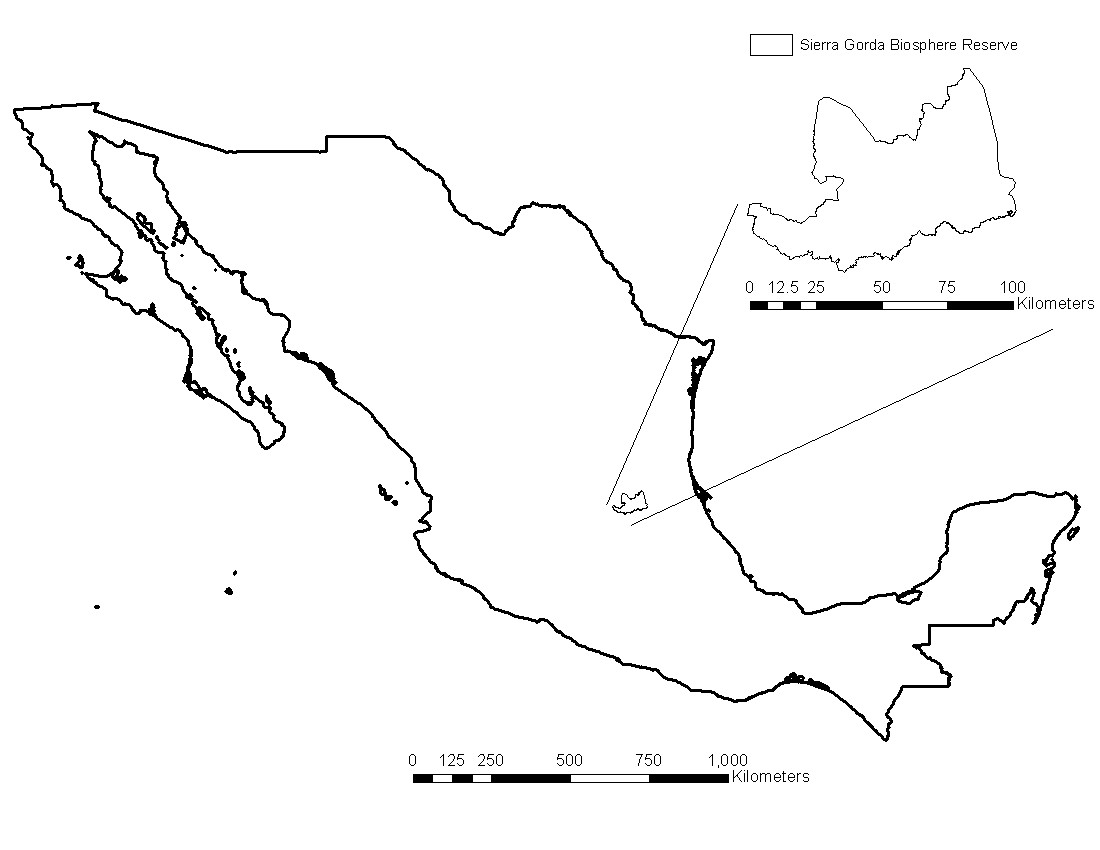
Figure 1. Location of the Sierra Gorda Biosphere reserve in central Mexico
In central Mexico, there is a geographically and biologically unique space featuring a mosaic of a varied diversity of flora and fauna, the Sierra Gorda Biosphere Reserve. Yet similar to other reserves in the world, human and climate drivers are changing the unique conditions of this area at an accelerated rate (Figure 2). However, this reserve remains home to important protected and endangered species that are struggling to find optimal habitat conditions elsewhere, thus it is important to identify potential areas that are suitable for keep these animal populations.Jaguar (Panthera onca) is one of these endangered species, and essential in the faunal community because of its position as a top predator (Figure 2).
Figure 2. Jaguar captured in a camera trap by Grupo Ecologico Sierra Gorda
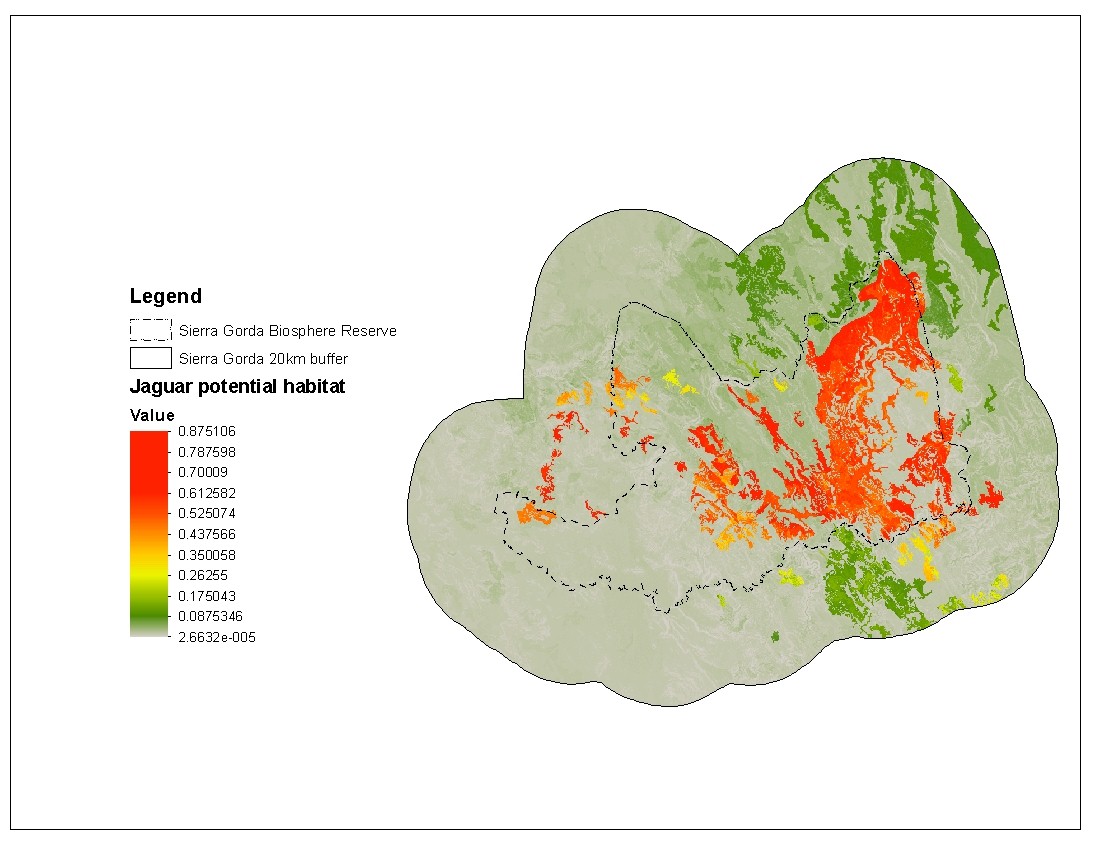
Furthermore, the presence of jaguars is an indicator of the health of the ecosystems and by conserving its habitat we can also protect other species. However, is very difficult to allocate efforts to large areas and therefore we need to identify and prioritize the most important zones capable to host jaguars.In the SILVIS lab, Carlos Ramirez Reyes, created a potential species distribution model for jaguars based on presence data and using factors than can affect their distribution such as topography, landcover and precipitation among others (Figure 3).
Figure 3. Potential distribution for jaguars in the Sierra Gorda Biosphere Reserve
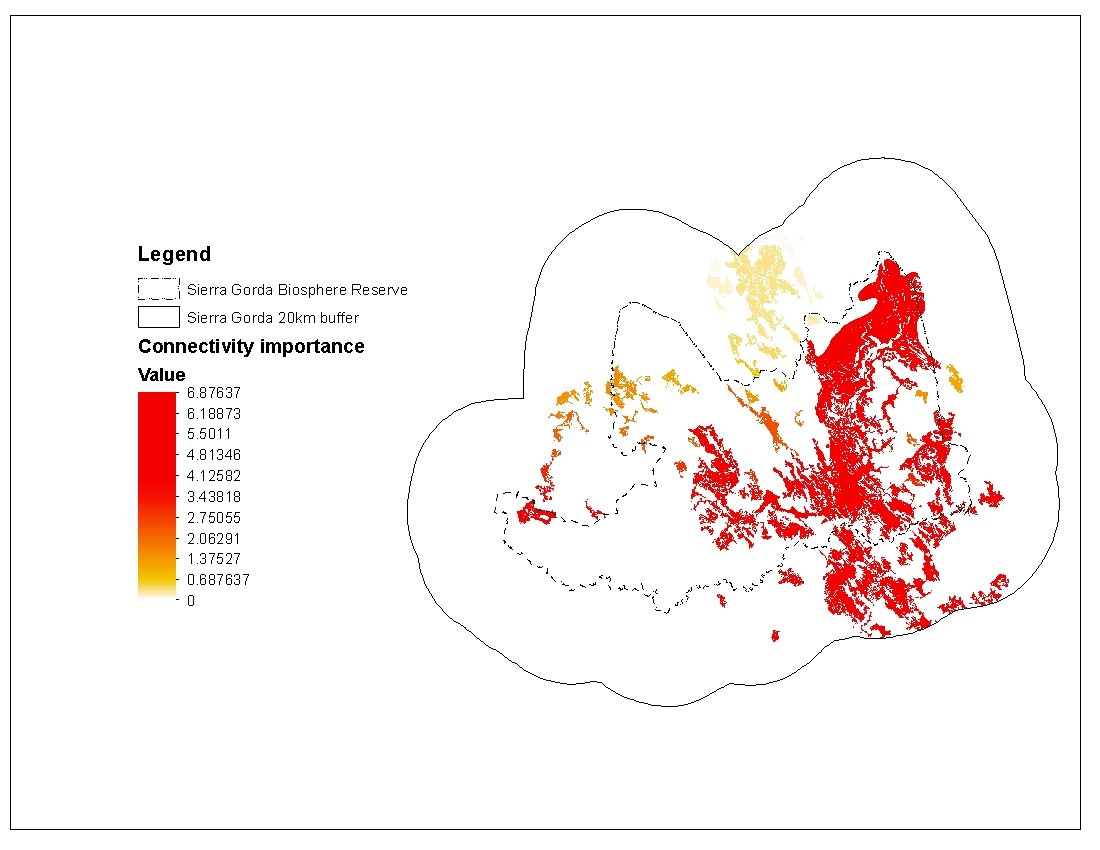
Based on this model, he identified areas inside within and outside the reserve that could host jaguars. Additionally, by adding connectivity parameters to the models (Figure 4), he evaluated if habitat connectivity actually improves the potential distribution model for the reserve. The goal was to find patches of potential habitat that are critical for connectivity of the entire reserve.
Figure 4. Relative importance of potential habitat patches to maintain the connectivity of the system
With his research Carlos hopes to gain better knowledge for the habitat requirements of the jaguar in the region. Ultimately, the project should serve to inform nongovernmental organizations and government agencies with interests in the reserve to make decisions on how to distribute resources for the management of the species and prioritize areas that should be protected. Furthermore, this project includes a new method for obtaining potential areas through the use of landscape connectivity. Similar projects that aim to model potential species distribution can benefit from this technique given that landscape connectivity is also important for animal dispersal and gene flow in fragmented landscapes. “
Conservation plans identify where we can most efficiently protect parcels across the landscape to help conserve biodiversity into the future. Plans are comprehensive, involving the efforts of many people, including the public, often over multiple years. As a result, plans can be costly to develop. Thus it’s important to understand whether plans are making a difference. That is, are conservation plans actually influencing how and where we protect lands.
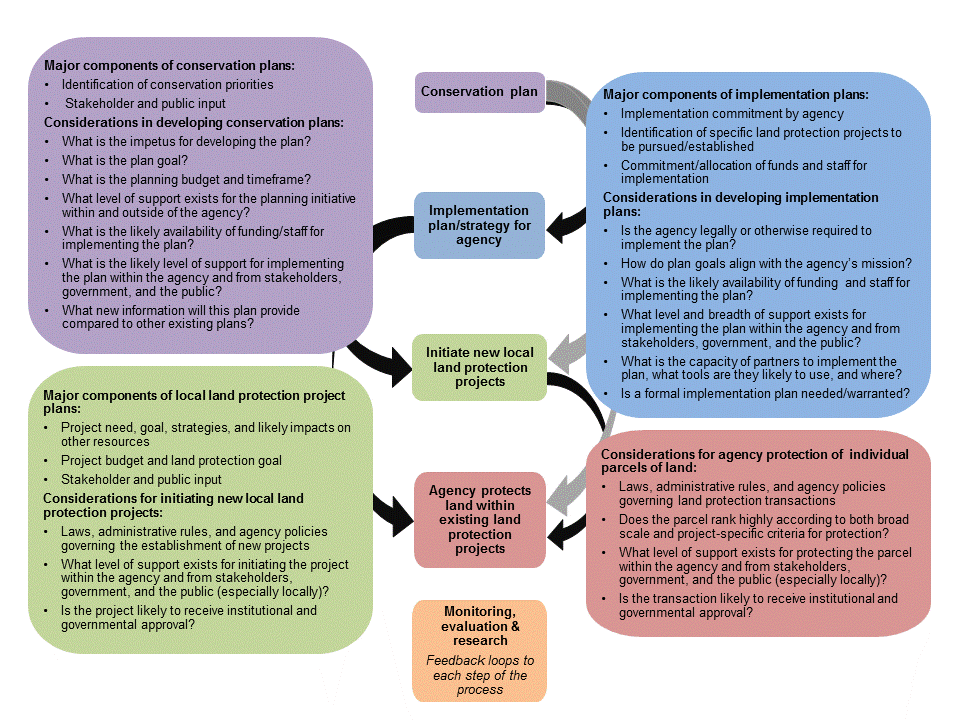
Figure 1. The comprehensive and interconnected nature of developing and implementing conservation plans.
Sarah looked at two different kinds of plans – broad-scale plans that set priorities for land protection across an entire state, and local-scale plans that help define exactly which parcels within a given priority area we seeking to protect. She asked whether each type of plan influenced 1) how much land was protected, 2) where land was protected, and 3) what type of land was protected (e.g., forests for grasslands). Sarah asked these questions in the state of Wisconsin, in part because the Wisconsin Department of Natural Resources has been actively planning for and conserving land in Wisconsin for more than a century. The actual impact of conservation plans is difficult to detect, because we don’t know what would have happened in the absence of a plan. History is tricky like that. So Sarah took a different approach. She looked at land protection actions before and after each plan was developed. In essence, she asked whether we could see a difference in how much, where, or what kinds of lands were protected before versus after each plan was completed. Sarah’s study focused on four statewide plans led or developed in close collaboration with the Wisconsin DNR. The plans were completed in 1939, 1964, 2004, and 2008. She also looked at hundreds of local Wisconsin DNR plans completed between 1938 and 2011. Including historical plans (and land protection records) in the study provided an uncommon opportunity to look at the potential influence of plans over long time periods.
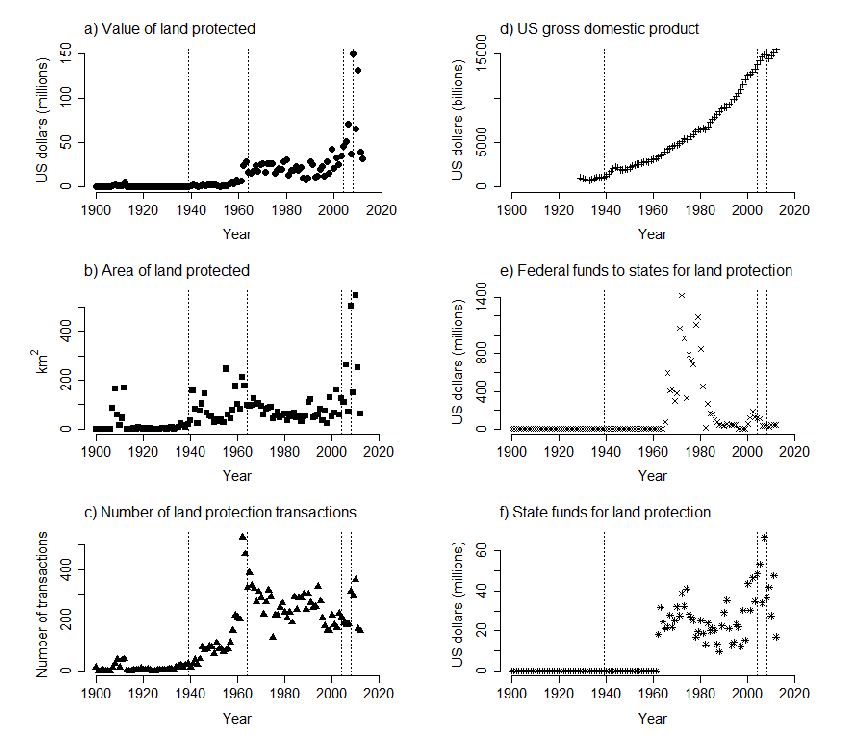
Fig. 2. Total annual statewide a) value of land protected, b) area of land protected, and c) number of land protection transactions; and d) US gross domestic product, e) federal funding granted to states for land protection, and f) state funds for land protection for each year between 1900 and 2012. Dotted lines indicate years in which conservation plans were completed
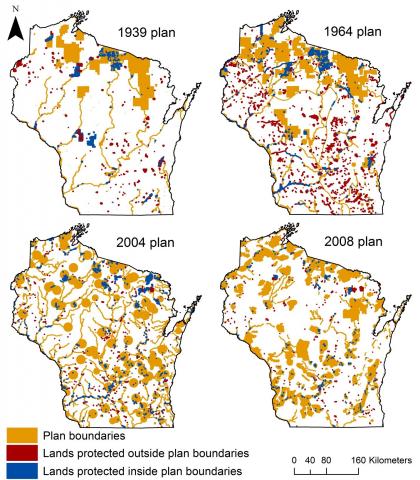
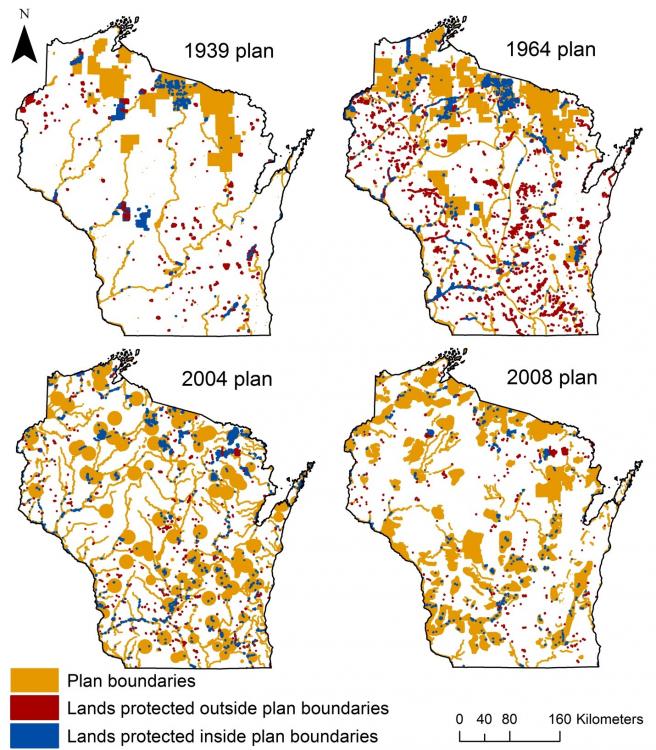
What Sarah found was in some ways surprising, and in other ways not. At a broad-scale, statewide plans did not have a clear influence on how much, where, or what type of lands are protected. Some changes in behavior that one might expect were detected. For example, the value and area of land protected in Wisconsin was greater after than before the 1939 plan. Similarly, more land was protected (via more individual transactions) after versus before the 2008 plan (Figure 2).However, overall 58% of lands protected within 20 years of plan completion were outside of plan boundaries (Figure 3).
Figure 3: The location of lands protected inside and outside of plan boundaries within twenty years after plan completion for each of the four statewide plans for Wisconsin. The size of small protected areas has been exaggerated slightly to more clearly show their spatial distribution.
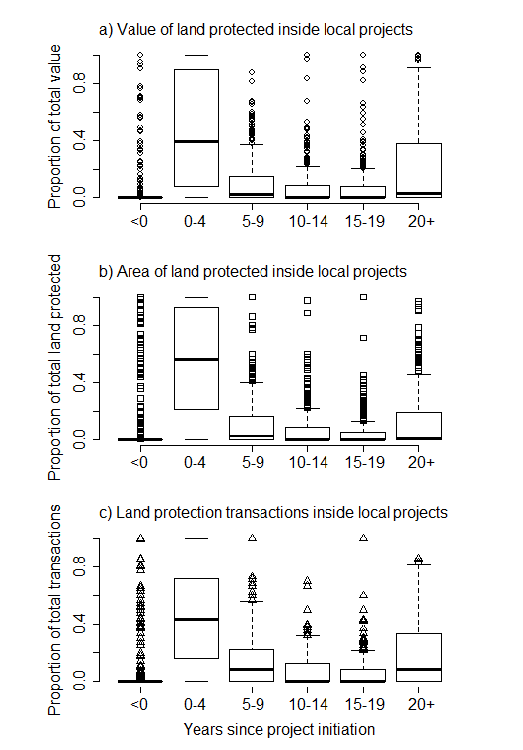
.There are a number of likely reasons for this. First, money matters. Often changes in land protection were associated with changes in funding for conservation, or in the economy in general. It’s not surprising that plans are more likely to have an influence on land protection when there is more funding available to protect land. Second, it’s clear that plans build on each other, making it difficult to separate out the influence of any individual plan. Thus priority areas in recent plans are often the same general areas that were identified in earlier plans. Conservation actions build on each other – areas that we protect improve in quality over time through management and restoration actions, and become even more valuable as core areas around which we can center future protection activities. Thus our recent actions are often a continuation of actions that began many years ago, decades before the most recent plan was written!…..The story was quite different when Sarah and her colleagues looked at local-scale plans (what the Wisconsin DNR calls local projects, or plans that cover individual parks or wildlife areas). More than 91% of the time, more activity occurred in a local area after a local plan was approved. Further, most of this activity happened within the first five years (Figure 4). Clearly local plans make a difference, and they do so in a hurry.
Fig. 4. Land protection activity over time inside individual, local land protection projects (n=371 projects).
The take home message: plans do matter. Broad-scale plans set the stage for local plans, and local plans correspond with big changes in local land protection. But clearly plans can be most influential when you have a perfect storm of influences coming together – when the economy is strong, when there is public and political support for conservation, and when there is funding available specifically for land protection for conservation purposes. While this conclusion will come as no surprise to many, it’s not something that had been documented previously in the scientific literature. The century-long, carefully-documented history of conservation planning and land protection by the Wisconsin DNR is not common worldwide, and allowed us to quantify and better understand this important interplay between plans, actions, and money in a way that hadn’t been possible before.”
In the northern region of Yunnan Province, for generations, Tibetan villagers have set aside patches of forest for spiritual proposes, also known as Tibetan sacred forests. During previous research Jodi and Eric discovered that sacred forests are keystone structures for conservation of forest bird in a landscape dominated by degraded pastureland.
Figure 1. Mrs. Gould’s Sunbird (Aethopyga gouldiae), which are often found using Tibetan sacred forests as breeding habitat. (Photo: Eric Wood)
They found more than 30 bird species heavily using sacred forests rather than pastureland. This result highlighted the importance of these forests for bird conservation. After this first study, the next step was to understand what characteristics of these forests make them so attractive for bird? Eric said: ‘In order to effectively manage and conserve bird species in the sacred forests, we need to understand the foundational habitat requirement of these birds. Once we know this, they we can begin to make recommendations for conservation applications. We now know that sacred forests support many bird species, the question that remain is why?’ For this, Jodi and Eric collected field data on habitat structure and composition and bird richness and abundance to construct bird-habitat relationship occupancy models. Some key structural variables that the pair sampled were cover of leaf litter, herbaceous materials, shrubs and trees. These variables are indicators of what species need for survivorship: nesting sites, food, refugee and protection. Once data was collected, and back in the comfort of Madison, Eric developed models to relate the explanatory variables with occupancy of over 30 bird species. With this model-selection approach, it was possible to select best-supported habitat-characteristics associated with a particular species occupancy.
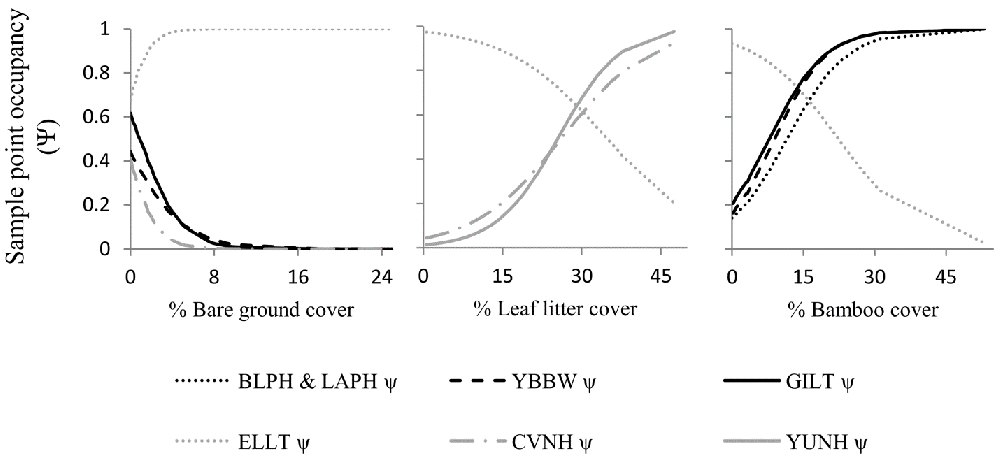
Figure 2 Relationships of predicted sample -point occupancy for six bird species with three habitat characteristics representing the ground and sub-canopy layer. Bird species codes: BLPH & LAPH = Blood and Lady Amherst’s Pheasant, YBBW = Yellowish-bellied Bush Warbler, GILT = Giant Laughingthrush, ELLT = Elliot’s Laughingthrush, CVNH = Chestnut-vented Nuthatch, YUNH = Yunnan Nuthatch.
In addition to the structural variables indicating diverse forested habitats, they found that high cover of leaf litter and shrubs are associated with occupancy of understory birds. While doing fieldwork, they observe that some understory specialist look for their food in the litter, where they can find insects underneath the leaves.
Figure 3. Gray-backed Shrike (Lanius tephronotus), which are often found using degraded habitat adjacent to Tibetan sacred forests. (Photo: Eric Wood)
These result highlighted the importance of field observation. In cases where so little life history information is available, field work become a crucial experience to understand what the models are saying to us afterwards. While China continues to grow in population, there is an increase pressure in resources (e.g., wood, water, and land) in less populated regions, as is the case of Yunnan. In addition to learning more about birds that use Tibetan sacred forests as breeding habitat, the result of this research helps inform future forest management in a regional context.”